گیرنده های جفت شده با پروتئین G:بررسی کامل
گیرنده های جفت شده با پروتئین G و مسیرهای پیامرسانی با انها
گیرنده های جفت شده با پروتئین G (GPCRs) خانواده ای از گیرنده های سطح سلولی هستند که نقش مهمی در بسیاری از فرآیندهای فیزیولوژیکی دارند. آنها در تنظیم ادراک حسی، انتشار انتقال دهنده های عصبی، ترشح هورمون و بسیاری از عملکردهای دیگر نقش دارند. در نتیجه، GPCR ها به یک هدف محبوب برای توسعه دارو تبدیل شده اند.
ما اصول اولیه سیگنال دهی GPCR، مسیرهای مختلف درگیر، و نقش پروتئین های G در این مسیرها را بررسی خواهیم کرد. بنابراین، چه دانشجوی فارماکولوژی یا یک متخصص بیوشیمی هستید، برای به دست آوردن درک عمیق تر از سیگنال دهی GPCR و پیامدهای آن برای توسعه دارو، ادامه مطلب را بخوانید.
مقدمه ای بر گیرنده های جفت شده با پروتئین G
گیرنده های جفت شده با پروتئین G یک کلاس جذاب از گیرنده های سطح سلولی هستند که نقش مهمی در سیگنال دهی سلولی دارند. آنها در طیف وسیعی از فرآیندهای بیولوژیکی از جمله ادراک حسی، انتقال عصبی، تنظیم هورمون و پاسخ ایمنی درگیر هستند. در واقع، GPCR ها یکی از بزرگترین خانواده های پروتئین در ژنوم انسان هستند که صدها گیرنده مختلف شناسایی شده است.
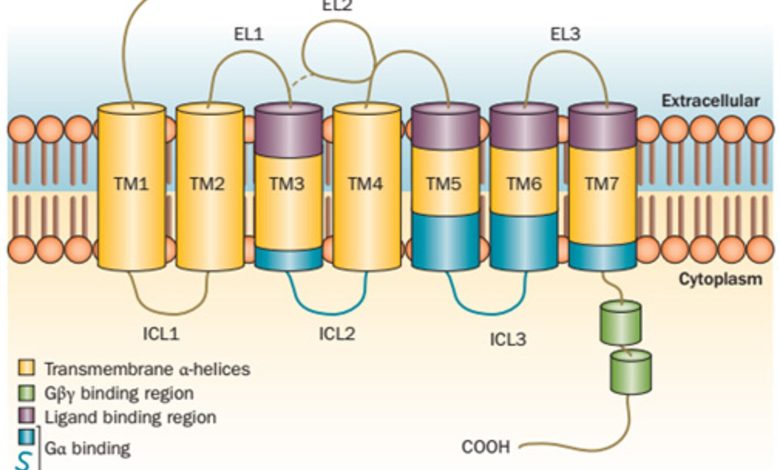
برای درک اهمیت GPCR ها، درک ساختار و عملکرد آنها ضروری است. این گیرنده ها از یک زنجیره پلی پپتیدی منفرد تشکیل شده اند که هفت بار غشای سلولی را می پوشاند و هفت حوزه گذرنده مارپیچ الفا را تشکیل می دهد. انتهای N گیرنده در خارج از سلول قرار دارد، در حالی که پایانه C در داخل سیتوپلاسم قرار دارد.
ویژگی کلیدی GPCR ها در توانایی آنها در تعامل با پروتئین های سیگنال دهی داخل سلولی به نام پروتئین های G نهفته است. پس از اتصال به لیگاندهای خاص خود مانند هورمون ها، انتقال دهنده های عصبی یا مولکول های حساس به نور، GPCR ها دچار تغییرات ساختاری می شوند که باعث فعال شدن پروتئین های G می شود. این فعال سازی، به نوبه خود، آبشاری از رویدادهای سیگنال دهی درون سلولی را آغاز می کند که در نهایت منجر به پاسخ های سلولی مختلف می شود.
مسیر های پیامرسانی گینده های جفت شده با پروتئین G
مسیرهای سیگنال دهی GPCR ها را می توان به دو نوع اصلی طبقه بندی کرد: مسیرهای جفت شده با پروتئین Gs و مسیرهای جفت شده با پروتئین Gi. پروتئین های Gs تولید آدنوزین مونوفسفات حلقوی (cAMP) را تحریک می کنند، در حالی که پروتئین های Gi مانع از تولید آن می شوند. این تعادل ظریف بین مسیرهای سیگنالینگ Gs و Gi، فرآیندهای سلولی مانند بیان ژن، فعالیت کانال یونی و فعال شدن آنزیم را تنظیم می کند.
علاوه بر این، GPCR ها همچنین می توانند سایر مسیرهای سیگنال دهی پایین دستی، مانند مسیر پروتئین کیناز فعال شده با میتوژن (MAPK) و مسیر phosphoinositide 3-kinase (PI3K) را فعال کنند تا پاسخ های سلولی خاصی را ایجاد کنند.
درک اصول اولیه سیگنال دهی سلولی
سیگنال دهی سلولی یک فرآیند پیچیده و پیچیده است که نقش مهمی در عملکردهای مختلف بیولوژیکی ایفا می کند. در قلب این فرآیند مسیرهای سیگنال دهی وجود دارد که ارتباط بین سلول ها را تسهیل می کند و پاسخ آنها به سیگنال های خارجی را هماهنگ می کند.
گیرنده های جفت شده با پروتئین G (GPCRs) یک کلاس برجسته از گیرنده های سطح سلولی هستند که در سیگنال دهی سلولی نقش دارند. این گیرنده ها در طیف وسیعی از فرآیندهای فیزیولوژیکی از جمله ادراک حسی، تنظیم هورمون و انتقال عصبی نقش دارند. درک اصول اولیه سیگنال دهی سلولی برای درک عملکرد پیچیده GPCR ضروری است.
به محض اتصال لیگاند، GPCRها دستخوش تغییرات ساختاری می شوند که پروتئین های سیگنال دهی داخل سلولی به نام پروتئین های G را فعال می کند. این پروتئینها بهعنوان سوئیچهای مولکولی عمل میکنند و بین حالت غیرفعال متصل به GDP و حالت فعال متصل به GTP تغییر میکنند. فعالسازی پروتئینهای G باعث بسیاری از رویدادهای سیگنالدهی پاییندستی میشود که میتواند شامل فعال شدن آنزیمها، مدولاسیون کانالهای یونی یا تنظیم بیان ژن باشد.
یکی از ویژگی های کلیدی GPCR ها توانایی آن ها در درگیر شدن با مسیرهای سیگنالینگ متعدد به طور همزمان است. این تطبیق پذیری به این گیرنده ها اجازه می دهد تا طیف گسترده ای از پاسخ های سلولی را واسطه کنند و تنظیم دقیق انتقال سیگنال را امکان پذیر می کند. تنوع مسیرهای سیگنال دهی GPCR با وجود زیرگروه های مختلف پروتئین G و عوامل مرتبط با آنها تقویت می شود. هر زیرگروه پروتئین G می تواند عوامل پایین دست متمایز را فعال کند، که منجر به پاسخ های سلولی منحصر به فرد می شود.
نقش GPCR ها در سیگنال دهی سلولی
مکانیسم سیگنال دهی GPCR با اتصال یک لیگاند خاص، مانند یک هورمون یا انتقال دهنده عصبی، به حوزه خارج سلولی گیرنده آغاز می شود. این رویداد اتصال باعث ایجاد یک تغییر ساختاری در گیرنده می شود که منجر به فعال شدن شرکای سیگنال دهی درون سلولی آن می شود.
پس از فعال شدن، GPCR ها با پروتئین های تخصصی معروف به پروتئین های G، که به عنوان سوئیچ های مولکولی عمل می کنند، تعامل دارند. این پروتئین های G از سه زیر واحد به نام های آلفا، بتا و گاما تشکیل شده اند. در حالت غیر فعال، زیر واحد آلفا به گوانوزین دی فسفات (GDP) متصل می شود. با این حال، پس از فعال شدن GPCR، تولید ناخالص داخلی با گوانوزین تری فسفات (GTP) مبادله می شود که منجر به جدا شدن زیرواحد آلفا از کمپلکس بتا گاما می شود.
زیرواحد آلفای آزاد شده، که اکنون به GTP متصل شده است، می تواند با پروتئین های موثر مختلف، مانند آنزیم ها یا کانال های یونی، برای شروع آبشارهای سیگنال دهی پایین دست تعامل کند. از سوی دیگر، کمپلکس بتا گاما همچنین می تواند مسیرهای سیگنالینگ را با تعامل مستقیم با سایر اجزای سلولی تعدیل کند.

نکته مهم، مدت و شدت سیگنال دهی GPCR به شدت تنظیم می شود. زیرواحد آلفای متصل به GTP دارای یک فعالیت GTPase ذاتی است که هیدرولیز GTP به GDP را کاتالیز می کند. این فرآیند منجر به غیرفعال شدن زیرواحد آلفا میشود و به آن اجازه میدهد تا با کمپلکس بتا-گاما ارتباط برقرار کند و سیگنالدهی را خاتمه دهد.
علاوه بر این، سیگنال دهی GPCR را می توان با مکانیسم های تنظیمی مختلف، از جمله حساسیت زدایی گیرنده و درونی سازی تعدیل کرد. حساسیت زدایی شامل فسفوریلاسیون گیرنده فعال شده توسط کینازهای خاص است که اتصال پروتئین های تنظیمی به نام آرستین را تقویت می کند. این فعل و انفعال از جفت شدن بیشتر پروتئین G جلوگیری می کند و باعث اندوسیتوز گیرنده می شود که در نهایت منجر به تضعیف سیگنالینگ می شود.
به طور خلاصه، گیرنده های جفت شده با پروتئین G به عنوان واسطه های مهم سیگنال دهی سلولی عمل می کنند و سیگنال های خارج سلولی را به پاسخ های درون سلولی منتقل می کنند. درک مسیرهای پیچیده درگیر در سیگنال دهی GPCR برای کشف پیچیدگی های ارتباطات سلولی و توسعه درمان های هدفمند برای بیماری های مختلف حیاتی است.
مروری بر مسیرهای سیگنال دهی GPCR
مسیرهای سیگنال دهی GPCR ها می تواند پیچیده و متنوع باشد، اما معمولاً از یک الگوی مشابه پیروی می کنند. هنگامی که یک لیگاند، مانند یک هورمون یا یک انتقال دهنده عصبی، به GPCR در سطح سلول متصل می شود، باعث ایجاد آبشاری از رویدادها می شود که در نهایت منجر به پاسخ سلولی می شود.
اولین مرحله در سیگنال دهی GPCR شامل فعال شدن پروتئین های هتروتریمری G است که از سه زیرواحد آلفا، بتا و گاما تشکیل شده اند. اتصال لیگاند به گیرنده باعث تغییر ساختاری می شود که به GPCR اجازه می دهد با پروتئین G تعامل داشته باشد. این تعامل منجر به تبادل GDP (گوانوزین دی فسفات) برای GTP (گوانوزین تری فسفات) در زیر واحد آلفا پروتئین G می شود که منجر به جدا شدن زیر واحد آلفا از کمپلکس بتا گاما می شود.
زیرواحد آلفای فعال شده که اکنون به GTP متصل شده است، قادر به تعامل با پروتئین های مؤثر مختلف مانند آدنیلیل سیکلاز، فسفولیپاز C یا کانال های یونی است. این پروتئینهای مؤثر، رویدادهای سیگنالدهی درون سلولی را واسطه میکنند که در نهایت منجر به پاسخ سلولی میشود. به عنوان مثال، فعال شدن آدنیلیل سیکلاز توسط زیرواحد آلفا پروتئین G منجر به تولید دومین AMP حلقوی پیام رسان (cAMP) می شود که می تواند فرآیندهای سلولی مختلف را تنظیم کند.
علاوه بر مسیرهای با واسطه پروتئین G، GPCR ها می توانند مسیرهای سیگنالینگ دیگری مانند مسیر بتا آرستین را نیز فعال کنند. بتا آرستین ها پروتئین هایی هستند که می توانند به GPCR فعال شده متصل شوند و منجر به درونی شدن و حساسیت زدایی آن شوند. با این حال، بتا آرستین ها همچنین می توانند به عنوان داربست های سیگنال، جذب پروتئین های دیگر و شروع آبشارهای سیگنالینگ جایگزین نیز عمل کنند.
اتصال لیگاند و فعال شدن گیرنده
در هسته سیگنال دهی GPCR برهمکنش بین یک مولکول لیگاند و گیرنده مربوطه آن است. لیگاندها می توانند متنوع باشند، از مولکول های کوچک مانند انتقال دهنده های عصبی، هورمون ها یا حتی فوتون ها در مورد GPCR های بینایی تا مولکول های بزرگتر مانند پپتیدها یا پروتئین ها. هنگامی که یک لیگاند به دامنه خارج سلولی GPCR متصل می شود، باعث ایجاد تغییر ساختاری در گیرنده می شود که منجر به فعال شدن آن می شود.
رویداد اتصال لیگاند باعث ایجاد آبشاری از رویدادهای سیگنال دهی پایین دستی از طریق GPCR می شود. در حالت غیر فعال خود، GPCR به یک پروتئین G هتروتریمری متشکل از سه زیر واحد – آلفا، بتا و گاما جفت می شود. پس از اتصال لیگاند، GPCR دچار یک تغییر ساختاری می شود که به آن اجازه می دهد با پروتئین G تعامل داشته باشد.
این تعامل منجر به تبادل GDP (گوانوزین دی فسفات) متصل به زیر واحد پروتئین G آلفا با GTP (گوانوزین تری فسفات) می شود و در نتیجه پروتئین G به زیر واحدهای آلفا و بتا گاما تجزیه می شود. هر دو زیرواحد آلفا و زیرواحد بتا گاما می توانند به طور مستقل مسیرهای سیگنال دهی پایین دستی مختلف، از جمله فعال سازی آنزیم ها یا کانال های یونی، تعدیل بیان ژن، و تنظیم پیام رسان های دوم درون سلولی را فعال کنند.
اتصال لیگاند و فعال شدن گیرنده متعاقب آن فرآیندهای بسیار پویا هستند که مدت و قدرت سیگنال به خوبی تنظیم می شود. پس از فعال شدن، GPCR تحت حساسیت زدایی قرار می گیرد، فرآیندی که از سیگنال دهی پایدار جلوگیری می کند. این می تواند از طریق مکانیسم هایی مانند فسفوریلاسیون گیرنده توسط کینازها اتفاق بیفتد که منجر به جذب آرستین ها می شود که از فعال شدن بیشتر پروتئین G جلوگیری می کند.
فعال شدن پروتئین های G
فعالسازی پروتئینهای G یک گام مهم در مسیرهای سیگنالدهی گیرندههای جفت شده با پروتئین G (GPCRs) است.
هنگامی که یک لیگاند، مانند یک هورمون یا انتقال دهنده عصبی، به GPCR متصل می شود، باعث ایجاد تغییر ساختاری در گیرنده می شود. این تغییر ساختاری به GPCR اجازه می دهد تا با پروتئین های G خاص واقع در سمت داخل سلولی غشای سلولی تعامل داشته باشد و آن را فعال کند.
پروتئین های G پروتئین های هتروتریمری هستند که از سه زیر واحد آلفا، بتا و گاما تشکیل شده اند. در حالت غیرفعال، زیر واحد آلفا به GDP (گوانوزین دی فسفات) متصل است و کمپلکس بتا گاما با زیر واحد آلفا مرتبط است.
پس از فعال شدن GPCR، گیرنده با پروتئین G تعامل می کند و باعث تبادل GDP برای GTP (گوانوزین تری فسفات) در زیر واحد آلفا می شود. این تبادل منجر به جداسازی زیرواحد آلفا از کمپلکس بتا گاما میشود و هر دو جزء اکنون میتوانند به طور مستقل با مولکولهای مؤثر پاییندست تعامل داشته باشند.
زیرواحد آلفای فعال شده، همراه با GTP متصل به آن، به عنوان یک سوئیچ مولکولی عمل می کند و باعث ایجاد آبشاری از رویدادهای درون سلولی می شود. این می تواند مستقیماً مولکول های مؤثر مختلف مانند آنزیم ها یا کانال های یونی را تنظیم کند یا می تواند فعالیت پیام رسان های ثانویه مانند AMP حلقوی (cAMP) یا یون های کلسیم را تعدیل کند.
از سوی دیگر، کمپلکس بتا گاما همچنین میتواند با تعامل با سایر پروتئینهای درون سلولی در انتقال سیگنال شرکت کند. این می تواند کانال های یونی، آنزیم ها یا حتی سایر GPCR ها را تحت تاثیر قرار دهد و منجر به تقویت یا مدولاسیون سیگنال اولیه شود.
نکته مهم این است که مدت زمان سیگنال دهی GPCR به شدت تنظیم می شود تا از پاسخ های سلولی مناسب اطمینان حاصل شود. زیرواحد آلفا دارای فعالیت ذاتی GTPase است که به آن امکان می دهد GTP را به GDP هیدرولیز کند و منجر به غیرفعال شدن پروتئین G شود. این فرآیند پیوند مجدد زیرواحد آلفا را با کمپلکس بتا گاما تسهیل میکند و پروتئین G را به حالت غیرفعال باز میگرداند.
آبشارهای سیگنال دهی داخل سلولی
زیرواحد های پروتئین G به عنوان یک سوئیچ مولکولی عمل می کند و باعث فعال شدن مولکول های مؤثر مختلف مانند آدنیلیل سیکلاز، فسفولیپاز C یا کانال های یونی می شود. این افکتورها به ترتیب پیام رسان های ثانویه درون سلولی از جمله آدنوزین مونوفسفات حلقوی (cAMP)، اینوزیتول تری فسفات (IP3) و یون های کلسیم را تولید می کنند.
پیام رسان های ثانویه به نوبه خود مولکول های سیگنال دهنده پایین دستی مانند پروتئین کینازها را فعال می کنند که پروتئین های هدف را فسفریله می کنند. فسفوریلاسیون به عنوان یک مکانیسم تنظیمی کلیدی عمل می کند که فعالیت پروتئین های سلولی متعددی از جمله آنزیم ها، فاکتورهای رونویسی و کانال های یونی را تعدیل می کند. این تنظیم با واسطه فسفوریلاسیون امکان کنترل دقیق پاسخ های سلولی را فراهم می کند و نتایج فیزیولوژیکی مناسب را تضمین می کند.
علاوه بر این، GPCR ها می توانند مسیرهای سیگنالینگ دیگری مانند مسیر پروتئین کیناز فعال شده با میتوژن (MAPK) را نیز فعال کنند. این مسیر نقش مهمی در تکثیر سلولی، تمایز و بقا دارد. فعال سازی MAPK شامل یک آبشار فسفوریلاسیون متوالی است که در آن یک سری از پروتئین کینازها فسفریله می شوند و یکدیگر را فعال می کنند که در نهایت منجر به فعال شدن فاکتورهای رونویسی و تغییرات بیان ژن می شود.

به طور کلی، آبشارهای سیگنال دهی درون سلولی که توسط GPCR ها ایجاد می شوند، فرآیندهای بسیار هماهنگ و کاملاً تنظیم شده هستند. آنها مولکول ها و مسیرهای سیگنال دهی مختلف را برای برانگیختن پاسخ های سلولی خاص ادغام می کنند. درک این مکانیسم های پیچیده سیگنالینگ نه تنها برای تحقیقات پایه ضروری است، بلکه دارای پتانسیل درمانی بسیار زیادی است، زیرا بسیاری از بیماری ها با مسیرهای سیگنالینگ GPCR نامنظم مرتبط هستند.
پیام رسان های ثانویه و نقش آنها در سیگنال دهی GPCR
در دنیای پیچیده گیرندههای جفت شده با پروتئین G (GPCRs)، پیامرسانهای ثانویه نقش مهمی در میانجیگری مسیرهای سیگنالینگ دارند. این مولکولهای کوچک بهعنوان واسطه عمل میکنند و پیام را از GPCR به عوامل پاییندستی منتقل میکنند و در نهایت منجر به پاسخ سلولی میشوند.
یکی از شناخته شده ترین پیام رسان های دوم، آدنوزین مونوفسفات حلقوی (cAMP) است. با فعال شدن یک GPCR، پروتئین G مرتبط با تغییر ساختاری، منجر به فعال شدن آدنیلیل سیکلاز می شود. این آنزیم تبدیل آدنوزین تری فسفات (ATP) به cAMP را کاتالیز می کند. سپس افزایش سطح cAMP باعث ایجاد آبشاری از رویدادهای درون سلولی می شود که پروتئین کیناز A (PKA) را فعال می کند و متعاقباً بر فرآیندهای سلولی مختلف مانند رونویسی ژن، تنظیم کانال یونی و متابولیسم تأثیر می گذارد.
پیام رسان ثانویه مهم دیگر اینوزیتول تری فسفات (IP3) است. با فعالسازی GPCR، فسفولیپاز C (PLC) فعال میشود که منجر به هیدرولیز فسفاتیدیلنوزیتول 4،5-بیسفسفات (PIP2) به IP3 و دیآسیلگلیسرول (DAG) میشود. IP3 سپس به گیرنده خود در شبکه آندوپلاسمی متصل می شود و منجر به آزاد شدن یون های کلسیم (Ca2+) از ذخایر داخل سلولی می شود. افزایش سطح کلسیم درون سلولی به عنوان سیگنالی برای فرآیندهای سلولی مختلف از جمله انقباض عضلانی، آزادسازی انتقال دهنده های عصبی و فعال شدن آنزیم عمل می کند.
علاوه بر این، کلسیم می تواند به عنوان پیام رسان دوم مستقل از IP3 عمل کند. GPCR ها می توانند مستقیماً کانال ها یا ناقل های کلسیم را فعال کنند که منجر به هجوم یا خروج یون های کلسیم می شود. این هجوم کلسیم می تواند باعث ایجاد آبشارهای مختلف سیگنالینگ پایین دستی شود و پروتئین های وابسته به کلسیم مانند کالمودولین و پروتئین کیناز C (PKC) را فعال کند که عملکردهای سلولی را بیشتر تنظیم می کند.
سایر پیام رسان های ثانویه درگیر در مسیرهای سیگنال دهی GPCR عبارتند از گوانوزین مونوفسفات حلقوی (cGMP)، دی اسیل گلیسرول (DAG) و اکسید نیتریک (NO). هر یک از این پیام رسان های دوم مسیرهای سیگنال دهی منحصر به فرد خود را دارند که با اثرگذارهای پایین دست خاصی در تعامل هستند و بر پاسخ های سلولی متنوع تأثیر می گذارند.
درک نقش پیام رسان های دوم در سیگنال دهی GPCR برای کشف پیچیدگی این مسیرها بسیار مهم است. با رمزگشایی از تعامل پیچیده بین GPCR ها، پروتئین های G و پیام رسان های دوم، محققان می توانند بینشی در مورد مکانیسم های اساسی سیگنال دهی سلولی به دست آورند و به طور بالقوه اهداف درمانی جدیدی را برای بیماری ها و اختلالات مختلف کشف کنند.